Yining Nan is a postgraduate student at School of Management, University of Science and Technology of China. Her research mainly focuses on behavioral operations management
Tengfei Nie is a Professor at School of Management, Shandong University. He received his Ph.D. degree from the University of Science and Technology of China in 2014. His research mainly focuses on supply chain management, game theory, behavioral operations management, crowdfunding, and co-creation
Advance selling activities based on e-commerce platforms have received much attention from consumers, which is a two-stage sales mode. However, many consumers have indicated that they are relatively happy to pay a deposit and feel more burdened at the final payment stage. That is, consumers feel differently at the two moments even though they know they will pay the same total amount for the product. This psychological behavior can be explained by mental accounting, which means the cognitive-computational process by which individuals or households code, evaluate, and record financial behavior. With the use of advance selling, this research has developed a game theoretical model to investigate how consumers’ mental accounting affects the optimal pricing and ordering decisions of supply chain members under wholesale price and revenue sharing contracts. The analysis shows that under wholesale price contracts, regardless of the optimal wholesale price set by the supplier, a portion of consumers will forgo the deposit paid, and the optimal order quantity for the retailer will always be equal to the consumers’ demand at the final payment stage. In exceptional cases, the optimal wholesale price may be equivalent to the retail price. The supplier’s optimal wholesale price increases monotonically with the consumer’s time coefficient for the price and decreases monotonically with the consumer’s time coefficient for product valuation. Furthermore, under a revenue sharing contract, there is a situation where the supplier’s optimal wholesale price is equal to cost. Additionally, a supplier’s optimal wholesale price exists such that none of the consumers will forgo the deposit they have paid. The study contributes to the understanding of mental accounting in advance selling and has implications for supply chain contract design.
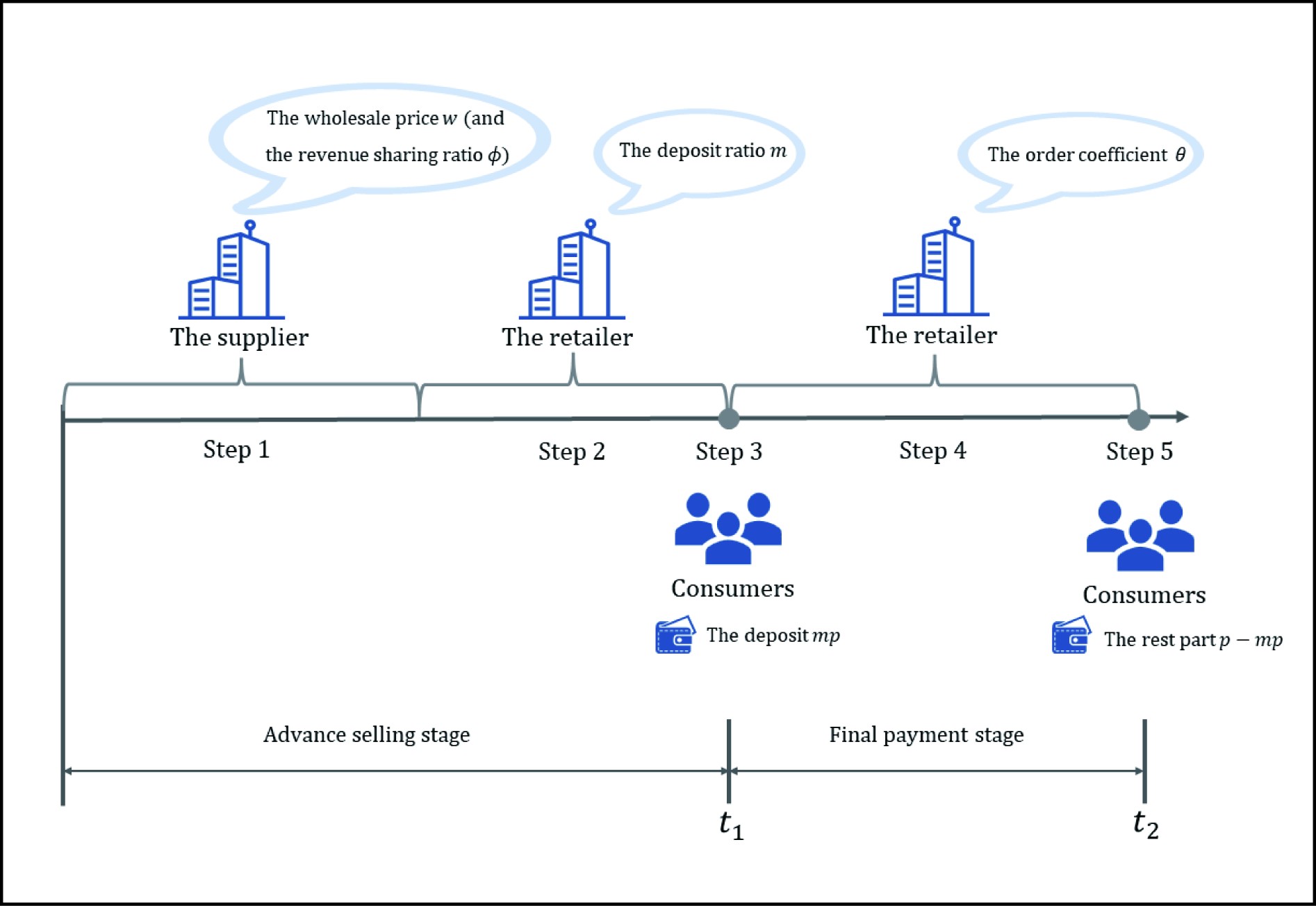
Graphical Abstract
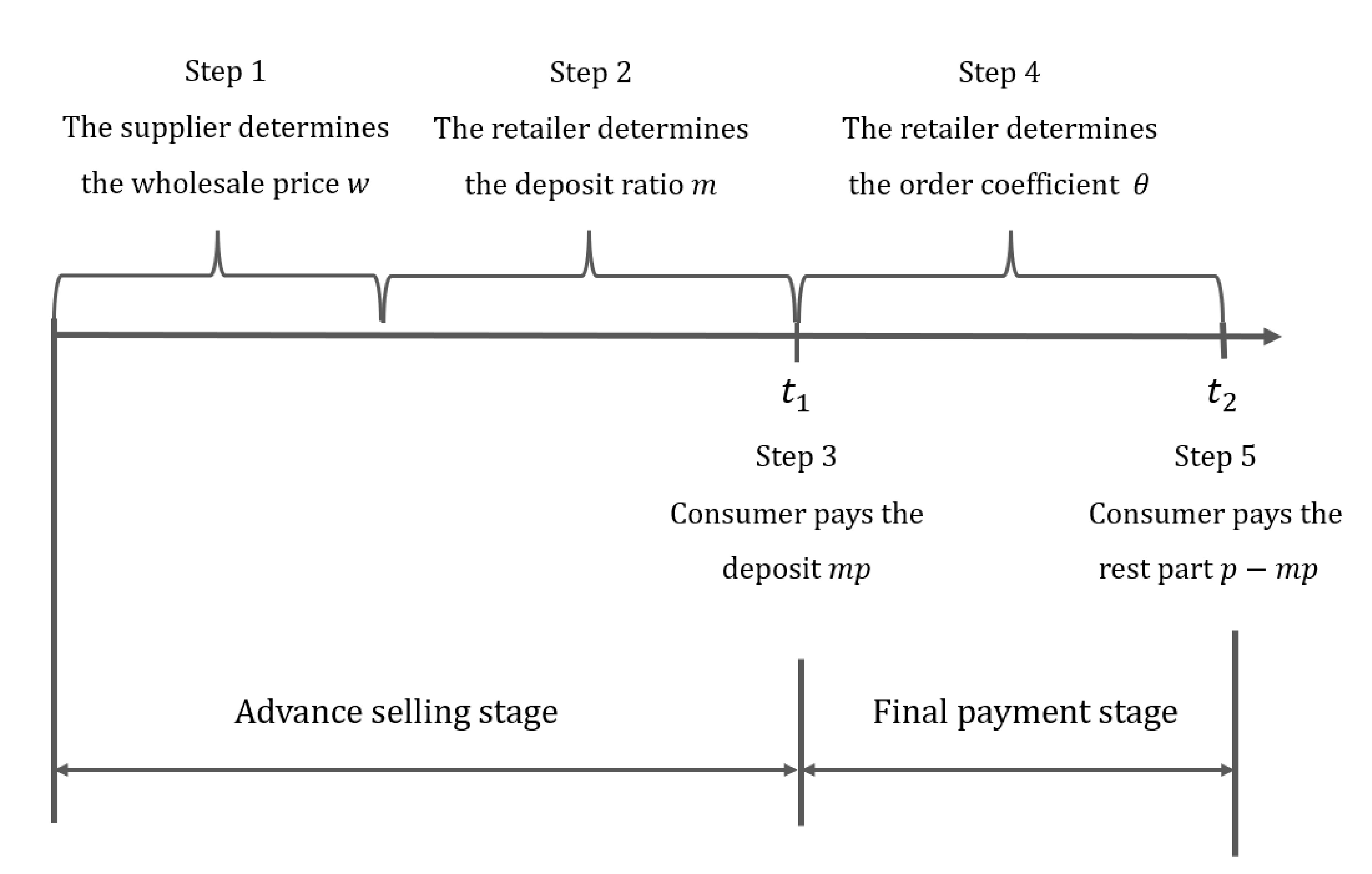
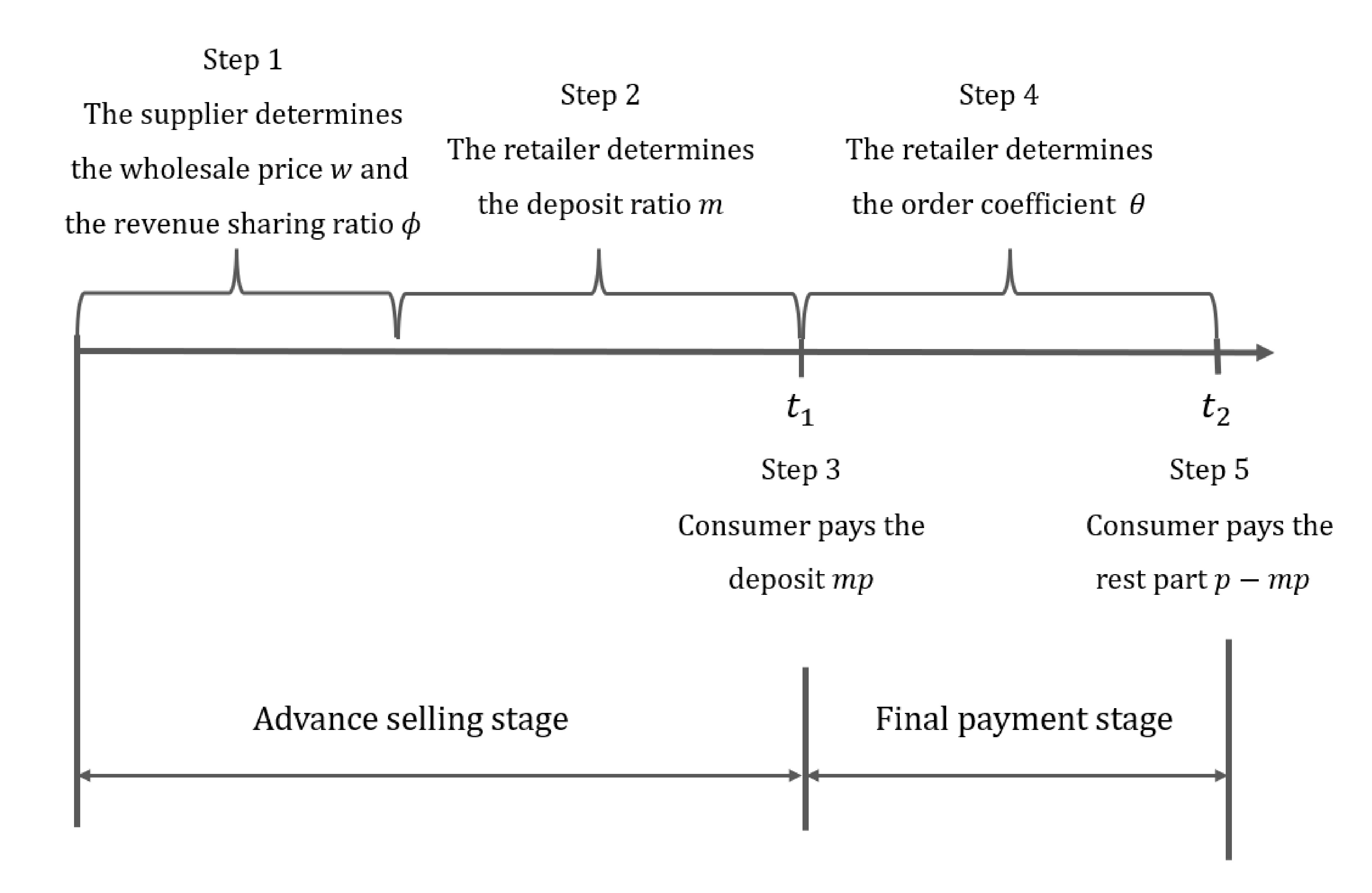
The structure of the paper containing the supply chain members’ decision sequence.
Abstract
Advance selling activities based on e-commerce platforms have received much attention from consumers, which is a two-stage sales mode. However, many consumers have indicated that they are relatively happy to pay a deposit and feel more burdened at the final payment stage. That is, consumers feel differently at the two moments even though they know they will pay the same total amount for the product. This psychological behavior can be explained by mental accounting, which means the cognitive-computational process by which individuals or households code, evaluate, and record financial behavior. With the use of advance selling, this research has developed a game theoretical model to investigate how consumers’ mental accounting affects the optimal pricing and ordering decisions of supply chain members under wholesale price and revenue sharing contracts. The analysis shows that under wholesale price contracts, regardless of the optimal wholesale price set by the supplier, a portion of consumers will forgo the deposit paid, and the optimal order quantity for the retailer will always be equal to the consumers’ demand at the final payment stage. In exceptional cases, the optimal wholesale price may be equivalent to the retail price. The supplier’s optimal wholesale price increases monotonically with the consumer’s time coefficient for the price and decreases monotonically with the consumer’s time coefficient for product valuation. Furthermore, under a revenue sharing contract, there is a situation where the supplier’s optimal wholesale price is equal to cost. Additionally, a supplier’s optimal wholesale price exists such that none of the consumers will forgo the deposit they have paid. The study contributes to the understanding of mental accounting in advance selling and has implications for supply chain contract design.
Public Summary
This paper extends the impact of consumer mental accounting on demand to the decisions of a supplier and a retailer, and applies it to wholesale price contracts and revenue sharing contracts.
This paper divides mental accounting by time dimension and portrays the role of mental accounting on consumers’ perception of product valuation and product price by two time coefficients.
This paper reveals the impact of mental accounting on consumers’ perception and purchase decisions, and draws some counterintuitive conclusions.
β-barrel outer membrane proteins (OMPs) are a defining feature of Gram-negative bacteria and play crucial roles in various cellular processes, including nutrient uptake, hazard shielding, and signaling. At the heart of their insertion into the outer membrane is BamA, a key component of the β-barrel Assembly Machinery (BAM) complex[1,2]. As a member of the Omp85 family, BamA is highly conserved across diderm bacteria, mitochondria, and chloroplasts[3]. In Escherichia coli (E. coli), it is one of two essential outer membrane proteins, primarily responsible for the biosynthesis of OMPs[4]. Exploring the mechanism by which BamA facilitates OMP folding is crucial for understanding membrane protein biogenesis and can provide implications for the development of antibiotics[5] that target bacterial pathogens.
Recently, significant strides have been made in elucidating the structure and function of the BAM complex, particularly in the model organism E. coli, primarily advanced by high-resolution cryo-electron microscopy (cryo-EM). The core protein BamA consists of two distinct regions: a C-terminal β-barrel domain, which is integrated into the outer membrane to facilitate the insertion of other OMPs, and a series of polypeptide transport-associated (POTRA) domains located at the N-terminus[6]. These POTRA domains, which are encircled by other BAM lipoproteins (BamB-E)[7], are instrumental in directing the β-signal of unfolded OMPs to the BAM complex. The structures of several E. coli BAM complexes, both with and without the OMP substrate, have been resolved, allowing researchers to characterize how EcBamA coordinates other BAM components and interacts with substrates to facilitate their folding and insertion process[8–12]: The periplasmic POTRA domains first recognize the C-terminal β-signal of the OMPs with the assistance of other BAM components, such as BamD. The unfolded OMP is subsequently transported to the β-barrel domain of BamA, which can open laterally and align with the substrate’s last β-strand at the C-terminus. The sequential insertion of other strands of the OMP into the membrane occurs while the first strand remains coupled with BamA’s β1 strand. This ‘open’ conformation of BamA is proposed to be a crucial intermediate state in the process of OMP folding. After complete folding and release of the OMP from the BAM complex into the hydrophobic outer membrane (OM) environment, the β1 and β16 strands of BamA reconverge into a ‘closed’ conformation.
Given that BamA is widely conserved, it is important to understand whether it has a unique catalytic mechanism across various bacteria. Bioinformatic studies have shown that while most Omp85 family proteins, including BamA, share a highly similar β-barrel domain, the POTRA domains display species-specific variations in sequence and number[13,14]. These variations in the POTRA domains are thought to facilitate the recognition of different substrate proteins across various bacterial species. Additionally, the BAM accessory lipoproteins (BamB, C, D, and E in E. coli), which interact with one or multiple POTRA domains, have also been found to vary and are not conserved among different bacteria[7,15,16]. Replacing the POTRA domains of EcBamA with those of other Gram-negative bacteria can cause cell growth defects and impede the OMP folding process[17,18]. The β-barrel domains from a range of bacteria, including Escherichia coli, Pseudomonas aeruginosa and Neisseria meningitidis, have been demonstrated to be functionally interchangeable[18], indicating that this domain operates via a conserved mechanism to fold OMPs, despite species differences. However, these orthologous BamA proteins predominantly originate from closely related proteobacteria species. It remains unknown whether the β-barrel domain from a distantly related bacterium could effectively replace its counterpart in E. coli.
Deinococcus radiodurans (D. radiodurans or Dr), a deep-branched diderm bacterium from the Deinococcota phylum (formerly Deinococcus-Thermus), which belongs to the Terrabacteria group, is thought to represent ancient evolutionary divergence[19]. Unlike its proteobacterial relatives, D. radiodurans possesses a distinctive outer membrane that lacks lipopolysaccharide (LPS)[20]. The BamA homolog DrBamA, encoded by the DR_0379 gene, is also pivotal for cell growth, division, and stability of the cell envelope, particularly under stress conditions[21]. In contrast to EcBamA from E. coli, which has five POTRA domains, DrBamA contains six POTRA domains. Furthermore, while E. coli has four lipoproteins (BamB to BamE) associated with BamA, no homolog has been identified in D. radiodurans. These marked differences make DrBamA an ideal candidate for testing the conservation of BamA’s function and understanding the roles of the β-barrel domain and the POTRA domains. In this study, we determined the near-atomic resolution structure of D. radiodurans BamA and demonstrated that the β-barrel domains in BamA are not interchangeable between D. radiodurans and E. coli. The distinct arrangements of BamA ECL4/6 among different bacterial species highlight the evolutionary diversity of BamA and provide new insights into the evolution of diderm bacteria.
2.
Materials and methods
2.1
Bacterial strains and growth conditions
DH5α[22] chemical competent cells (Alpalifebio) were used for plasmid subcloning. BL21 (DE3)[23] cells (Alpalifebio) were utilized for the expression and purification of DrBamA. All other strains used for functional tests were derived from the E. coli BW25113 strain[24]. Lysogeny broth (LB) and agar were prepared as previously described[25]. Unless otherwise specified, all liquid cultures were aerobically grown at 37 °C. When necessary, kanamycin (50 μg/mL), carbenicillin (50 μg/mL), streptomycin (50 μg/mL) and chloramphenicol (75 μg/mL) were added to the broth or agar. The strains are listed in Table S1 (see Supporting information).
2.2
Construction of plasmids
The dsDNA fragment encoding the signal peptide region (Met1–Ala21) and the functional region (Gln22–Trp846) of DrBamA were separately amplified from the genomic DNA of Deinococcus radiodurans R1[26] using primers oZW1–oZW2 and oZW3–oZW4 via polymerase chain reaction (PCR), respectively (Table S1). The gene encoding the tandem Flag-tag and His10-tag was amplified using primers oZW5 and oZW6. The gene encoding EcBamA was amplified from the genomic DNA of the BW25113 strain using primers oJC38 and oJC39. Phanta Max Super-Fidelity DNA Polymerase (Vazyme, P505-d1) was employed for the PCR amplification.
pZW01–02. To express DrBamA in E. coli, the native signal peptide (Met1–Ala21) was substituted with the 22-amino-acid pelB leader peptide from pectate lyase B in Erwinia carotovora, which is frequently used in the heterologous expression of outer membrane proteins in E. coli[27] and guides DrBamA to the outer membrane. Two affinity tags (Flag-tag and His10-tag) were inserted between pelB and DrBamA (Gln22–Trp846) to generate the pelB-tag-DrBamA construct (Fig. S1b). The gene encoding pelB-tag-DrBamA was cloned and inserted into the pCDFDuet[28] vector. The cloning process involved two steps. First, three dsDNA fragments encoding DrBamA (Met1–Ala21), Flag-His10, and DrBamA (Gln22–Trp846) were ligated and amplified via overlap PCR using primers oZW1 and oZW4. Simultaneously, the pCDFDuet vector backbone was amplified using primers oZW7 and oZW8. These two fragments were then conjugated to generate the plasmid pZW01 using the ClonExpress II One Step Cloning Kit (Vazyme, C112-01). Second, the pelB sequence was amplified from the pSB-13 plasmid (lab stock) using primers oZW9 and oZW10. The dsDNA encoding Flag-His10-DrBamA (Gln22–Trp846) was amplified from pZW01 using primers oZW5 and oZW8. The final plasmid, pelB-tag-DrBamA (pZW02), was created by ligating these two fragments together using the ClonExpress II One Step Cloning Kit (Vazyme).
pJC06. To enable arabinose-inducible expression of wild-type EcBamA, the bamA gene from E. coli was amplified and ligated with the pBAD33[29] vector backbone, which was PCR-amplified using primers oSY9 and oSY10, employing the ClonExpress II One Step Cloning Kit.
pZW23–25. To enable isopropyl β-D-thiogalactoside (IPTG)-inducible expression of EcBamA and DrBamA in E. coli, pZW24 and pZW25 were constructed on the basis of a plasmid bearing the T5lac promoter (pYD48, laboratory stock) using primers oZW29 and oZW35. For pZW24, where the wild-type EcBamA was fused with a Flag-tag (for immunoblotting assays), two dsDNA fragments encoding the EcBamA signal peptide (Met1–Gly20) and Flag-EcBamA (Ala21–Trp810) were initially amplified from pJC06 using primers oZW31–34. These fragments were subsequently ligated with the backbone of pYD48 using the ClonExpress II One-Step Cloning Kit. For pZW25, two dsDNA fragments encoding EcBamA (Met1–Gly20) and Flag-DrBamA (Gln22–Trp846) were amplified from pJC06 and pZW01 using primers oZW31–32 and oZW37–38 and ligated in the same manner. The empty vector pZW23 was constructed by amplifying and religating the backbone of the pYD48 plasmid using primers oZW29 and oZW35.
pZW26–29. Plasmids expressing chimeric EcBamA variants were generated from pZW01 and pZW03. Fragments of DrBamA (DrB, DrP, Dr4 and Dr6) were amplified from pZW01, while the fragments containing the truncated EcBamA gene with the plasmid backbone were amplified from pZW23. The corresponding primers are listed in Table S1.
pZW30–34. Plasmids expressing truncated or mutated EcBamA variants were generated from pZW23 through site-specific mutagenesis using Phanta Max Super-Fidelity DNA Polymerase. The primers used are listed in Table S1.
2.3
Construction of the BamA depletion strain
BW25113 wild-type E. coli cells were first transformed with plasmids expressing phage λ-Red recombinase (pKD46[29]) and arabinose-inducible EcBamA (pJC06). The kanamycin resistance cassette (kan cassette) was PCR-amplified from pKD13[29] using primers oJC40 and oJC41, which flanked a 60-nt homologous sequence of the region adjacent to bamA. This kan cassette was then introduced into BW25113 cells harboring pKD46 and pJC06 to replace the native bamA gene via electroporation. Candidates of the BamA depletion strain were selected on LB plates supplemented with kanamycin, chloramphenicol, and 0.2% (w/v) L-arabinose and verified via colony-PCR and Sanger sequencing. The loss of the temperature-sensitive pKD46 plasmid was confirmed via colony-PCR and plating assays. The generated BamA depletion strain JC20 requires 0.2% (w/v) L-arabinose to induce BamA expression from pJC06 to support its growth. Other plasmids expressing EcBamA variants were introduced into strain JC20 by electroporation and grown in the presence of 0.2% L-arabinose but without IPTG.
2.4
Protein expression and purification
E. coli BL21 (DE3) was transformed with the plasmid pZW02 to express DrBamA and subsequently cultured in LB medium supplemented with streptomycin at 37 °C until the OD600 reached 0.8. DrBamA expression was induced by 0.2 mM IPTG at 37 °C for 3 h. For DrBamA purification, the cells were collected and then resuspended in lysis buffer (25 mM Tris-HCl, 150 mM NaCl, pH 8.0) supplemented with 1 mM phenylmethanesulfonylfluoride (PMSF). After sonication, the cell debris was removed via centrifugation (8000 × g, 10 min, 4 °C). The supernatant was then ultracentrifuged (100000 × g, 1 h, 4 °C) to harvest the membrane pellet. The membrane pellet was then resuspended in lysis buffer, followed by the addition of 1% (w/v) lauryl maltose neopentyl glycol (LMNG) to extract membrane protein at 4 °C for 2 h. After an additional centrifugation step (23280 × g, 1 h, 4 °C), the supernatant containing solubilized membrane proteins was applied to an anti-Flag M2 affinity gel (Sigma). The resin was washed with buffer A (25 mM Tris-HCl, 150 mM NaCl, 0.005% glyco-diosgenin (GDN), pH 8.0) and eluted with buffer A supplemented with 0.2 mg/mL Flag peptide. The eluate was further purified via size-exclusion chromatography with a Superose 6 increase 10/300 column in buffer A. The main peak fractions were collected and concentrated to approximately 4 mg/mL for cryo-EM sample preparation.
2.5
Cryo-EM sample preparation and data acquisition
To prepare the cryo-EM samples for structural analysis, 3.5 μL of purified DrBamA was applied twice to glow-discharged copper grids (Quantifoil R 1.2/1.3). The grid was subsequently blotted for 3.5 s in a chamber set at 100% humidity and 8 °C and then flash-frozen in liquid ethane cooled with liquid nitrogen via a Vitrobot Mark IV (Thermo Fisher Scientific). The grid was loaded onto a Titan Krios electron microscope (FEI, Thermo Fisher Scientific) operated at 300 kV and equipped with a BioQuantum energy filter and a K3 direct electron detector (Gatan). Images were automatically acquired via EPU in superresolution mode at a nominal magnification of 81000 × g. The defocus values ranged from −1.6 to −1.8 μm. Each micrograph had an exposure time of 3.8 s, resulting in a total dose rate of approximately 50 e−/Å2. The image stacks were motion-corrected via MotionCor2 and binned twofold, resulting in a pixel size of 1.07 Å/pixel. Moreover, dose weighting was performed[30]. Defocus values were estimated via CTFFIND4[31].
2.6
Image processing
A complete dataset comprising 4038 micrographs was collected for DrBamA, from which a total of 5755354 bin2 particles were extracted via the template picker in cryoSPARC[32]. A partial dataset with 851 micrographs was selected to generate an initial 3D model. This initial model served as a reference for the selection of high-quality particles from the full dataset through Heterogeneous Refinement followed by Ab-initio Reconstructions. This process yielded a total of 728678 selected particles. These high-quality particles were employed as seeds for seed-facilitated classification[33], resulting in the identification of 1920510 good particles for the full dataset. The bin1 particles were subsequently updated. The bin1 particles selected through seed-facilitated classification underwent a single round of Heterogeneous Refinement followed by three rounds of Ab-initio Reconstruction in C1 symmetry. This led to the selection of 637638 high-quality particles and the generation of a 3D model at a resolution of 4.0 Å. These particles were further subjected to one round of Ab-initio Reconstruction in C2 symmetry, with a total of 607784 particles selected, resulting in a 3D reconstruction at 3.8 Å resolution after Non-uniform Refinement. Resolutions were estimated with the gold-standard Fourier shell correlation 0.143 criterion with high-resolution noise substitution[34,35]. The local resolution map was calculated via the “Local Resolution Estimation” tool in cryoSPARC[32].
2.7
Model building and refinement
The 3.8 Å map of DrBamA was used for model building. The model was built in Coot[36], with the structure of DrBamA predicted by Alphafold2[37] used as the initial model, which was fitted into the map in ChimeraX[38]. Real-space model refinement with secondary structure and geometry restraints was conducted in PHENIX[39]. Figures showing the structure were prepared with PyMOL[40] and ChimeraX[38].
2.8
Multiple sequence alignment
The protein sequences of BamAs from different species were downloaded from the UniProt Consortium[41]. The sequences of their barrel domains were subsequently used for multiple sequence alignment via the ClustalW tool in MEGA11[42]. The phylogenetic tree in Fig. 4 was obtained from Ref. [19] and represented via iTOL v6 (https://itol.embl.de/).
Figure
4.
Mapping of ECL4 and ECL6 features on bacterial phylogeny. This figure presents a phylogenetic tree based on Witwinowski et al.[19], collapsed at the phylum level via iTOL v6 (https://itol.embl.de/). It includes one or two BamA structures from each phylum, sourced from previous studies, this research, or AlphaFold2[45] predictions. ECL4 is categorized as either possessing an outer helix or not, with those containing an outer helix further classified on the basis of polar interactions with other extracellular loops. ECL6 classification hinges on the presence of a stem region. The detailed species names and corresponding UniProt accession numbers are available in Table S3.
The structure of the barrel domain of DrBamA was resolved in this study. The structures of the barrel domains of EcBamA (PDB code: 5D0O)[8] and StBamA (PDB code: 5OR1)[46] were obtained from RSCB PDB[43,44]; the others were obtained from the AlphaFold protein structure database[45] or were predicted at a local server employing AlphaFold 2[45]. Structure alignment was conducted in PyMOL[40].
2.10
Spot assay of EcBamA mutants
The BamA depletion strain, carrying different IPTG-inducible BamA variants, was initially cultured in LB media supplemented with kanamycin, chloramphenicol, carbenicillin, and 0.2% (w/v) arabinose to induce wild-type EcBamA overnight at 37 °C. The overnight cultures were then diluted 1∶100 into fresh LB media supplemented with the same additives and allowed to reach the exponential growth phase at 37 °C. The cultures were subsequently further diluted to an OD600 of 0.5 and then subjected to a tenfold serial dilution from 100 to 10−5 in fresh LB medium. A 1 μL aliquot of each dilution was spotted onto LB plates containing 100 μM IPTG, but without arabinose, to selectively express the specific BamA variant. The plates were incubated at 37 °C for 15–20 h and photographed.
2.11
Western blot
Western blotting was performed to examine the protein expression levels of BamA variants in the membrane. Five hundred milliliters of JC20 cells transformed with BamA variants were cultured to the exponential growth phase as described in Section 2.10. The BamA variants were subsequently induced with 0.1 mM IPTG for 1.5 h at 37 °C. Then, the cells were collected and resuspended in approximately 25 mL of lysis buffer and sonicated. The cell debris and unbroken cells were removed by centrifugation at 8000 × g for 10 min. The supernatant was centrifuged at 100000 × g for 60 min, and the membrane pellet was collected. The membrane pellet was suspended in lysis buffer and then solubilized with LMNG for 1 h at 4 °C. After centrifugation (23800 × g, 1 h, 4 °C), the supernatant was normalized by the Bradford (Bio-red) to a total concentration of protein at 4 mg/mL. The samples were mixed with 5×SDS–PAGE loading buffer and then heated for 20 min at 95 °C. The samples were subjected to SDS–PAGE and transferred to PVDF membranes. The PVDF membranes were blocked with nonfat milk (5%) for 1 h at room temperature and then incubated with anti-Flag (for BamA variants) and anti-FtsI (for FtsI) antibodies. The PVDF membranes were washed with TBST 3 times and then incubated with the corresponding secondary antibodies. The PVDF membranes were washed with TBST 3 times and then detected.
3.
Results
3.1
The β-barrel domain of BamA is not always interchangeable
To increase our understanding of the function of BamA across different bacterial species, we compared the structural intricacies of EcBamA[8] with those of five proteobacterial BamAs, referencing data from the RCSB PDB[43,44] and the AlphaFold Database[45]. The analysis, which focused on β-barrel domains, revealed striking similarities, with minimal divergence in the root mean square deviation (RMSD) values, which were all less than 1.1 Å (Fig. 1a). Multiple sequence alignment (MSA) revealed lower sequence similarity of DrBamA’s barrel domain to that of proteobacteria BamAs (Fig. 1b, Fig. S1a), suggesting relatively large differences between their barrel domains. Interestingly, the extracellular loops of DrBamA are particularly different.
Figure
1.
Chimeric EcBamA carrying the β-barrel domain of DrBamA cannot rescue the loss of native EcBamA. (a) Comparison of the barrel domain of BamA orthologs from several gamma- and beta-proteobacteria. Ec: E. coli, St: Salmonella enterica serovar Typhimurium, Hi: Haemophilus influenzae, Pm: Pasteurella multocida, Pa: Pseudomonas aeruginosa, Nm: Neisseria meningitidis. The structures of the barrel domains of EcBamA (PDB code: 5D0O)[8] and StBamA (PDB code: 5OR1)[46] were obtained from RSCB PDB[43, 44], and the others were obtained from the AlphaFold Protein Structure Database[45]. (b) Sequence alignment of the barrel domain of BamA orthologs from the species in (a). ECL4 and ECL6 of EcBamA were labeled. (c) Schematic diagram of the BamA depletion strain. (d) Spot assay of full-length DrBamA and the chimera BamA carrying different domains of DrBamA and EcBamA. EV: empty vector. The results are representative of at least three independent experiments. The same applies hereinafter.
We first created an E. coli BamA depletion strain (JC20), with the native bamA gene replaced by a kanamycin resistance cassette and complemented with wild-type EcBamA on an L-arabinose-inducible plasmid (pJC06) (Fig. 1c). We then introduced DrBamA, or its specific domains, via a separate IPTG-inducible plasmid (Fig. 1c). The results of the spot assay revealed that full-length DrBamA expression was lethal, even worse than that of the empty vector (EV) control (Fig. 1d, lines 1–3), suggesting some type of dominant negative effect of the heterogeneity of DrBamA in E. coli. Since the lipoprotein components and the number of BamA POTRA domains in D. radiodurans are different from those in E. coli, this difference is likely due to the inability of DrBamA POTRA domains to recognize the β-signals of E. coli OMPs. This finding aligns with previous studies on full-length BamA replacements[17]. The minimum growth of the empty vector control (100 dilution) is likely attributable to the basal level expression of wild-type BamA from the pBAD vector. Accordingly, replacing the POTRA domains of EcBamA (Ala21–Thr423) with DrBamA (Gln22–Gly480, DrP) also caused severe cell death, similar to full-length replacement (Fig. 1d, line 4). Furthermore, we explored the interchangeability of the β-barrel domains between EcBamA and DrBamA by substituting the β-barrel domains of EcBamA (Gly424–Trp810) with those of DrBamA (Ile481–Trp846, referred to as DrB). Interestingly, E. coli cells expressing EcBamA (DrB) also presented severe growth deficiency in terms of cell growth (Fig. 1d, line 5), indicating that the β-barrel domain is not universally interchangeable, especially when the sequence similarity diverges significantly (Fig. S1a). The western blot results confirmed that DrBamA and the chimeric EcBamAs were expressed and folded in E. coli (Fig. S3a), suggesting that these proteins are dysfunctional in facilitating the folding of other OMPs in this bacterium.
3.2
Structural insights into the DrBamA β-barrel domain through cryo-EM analysis
To further investigate why the EcBamA (DrB) chimera fails to function in E. coli cells, unlike other proteobacterial BamA orthologs, we embarked on a detailed structural analysis of the DrBamA β-barrel domain (Fig. 2a) via cryo-EM single-particle analysis. We first modified the signal peptide of DrBamA to pelB to ensure heterogeneous expression and proper folding within the E. coli outer membrane (Fig. S1b). After detergent screening, DrBamA was extracted with LMNG and purified in the presence of GDN micelles, resulting in well-behaved proteins. Then, cryo-EM samples were prepared, and a dataset with 4038 movies was collected for structural determination (Fig. S1c). The initial 2D classification revealed that DrBamA predominantly exists in a dimeric form (Fig. S1d). Finally, the cryo-EM map of DrBamA was determined at 3.8 Å resolution using 607784 selected particles with C2 symmetry, revealing the intricacies of the barrel domain (Fig. 2b–c, Fig. S1e, and Table S2). Notably, all six POTRA domains were absent from the structure, implying their inherent flexibility—this aligns with findings from previous studies, such as that on Salmonella enterica serovar Typhimurium BamA[46].
Figure
2.
Overall structure of the DrBamA dimer. (a) Topologic model of the DrBamA barrel domain. β-strands are shown as arrows, and the α-helix is shown as a cylinder. The transmembrane sheets, ECL4 and ECL6, are colored yellow orange, slate and deep purple, respectively. (b) Cryo-EM map of dimeric DrBamA with each protomer in individual colors. (c) Cartoon representation of the DrBamA dimer from the side view and top view, with one protomer colored rainbow and the other colored as in (a). (d) Polar interactions between the two protomers of DrBamA. (e) Comparison of the barrel domains of DrBamA and EcBamA (PDB: 5D0O)[8].
A detailed dimeric model was constructed on the basis of the 3.8 Å cryo-EM map, of which 369 side chains (Arg478–Trp846) were assigned to each protomer (Fig. S1, Fig. S2). Each protomer has a classic 16 transmembrane β-strands architecture with prominent extracellular loops, such as ECL4 and ECL6. The size and shape of the barrel are comparable to those of EcBamA (40 Å in length, 24 Å in width, and 32 Å in height, Fig. 2c–e)[8]. Moreover, both protomers were observed in the ‘closed’ conformation[8]. Two polar interactions between the two protomers, Arg478–Gln816 and Tyr502–Gly813, were observed in the structure (Fig. 2d).
Furthermore, the significant increase in the root mean square deviation (RMSD) to 2.5 Å compared with that of EcBamA (PDB: 5D0O)[8] underscores a distinct structural divergence, especially in the altered shape, position, and orientation of ECL4 and ECL6 (Fig. 2e). These structural discrepancies may contribute to the functional deficiencies observed in the EcBamA (DrB) chimera.
3.3
The structural and functional roles of ECL4 and ECL6 in EcBamA and DrBamA
To determine whether variations in the ECL4 and ECL6 structures of BamA affect its function, we meticulously compared these extracellular loops between DrBamA and proteobacterial BamAs. All the BamAs exhibit an α-helix with 7–12 residues (hereafter the outer helix) within ECL4 (Fig. 3a), albeit with DrBamA’s helix being more peripherally positioned and tilted (Fig. 2e). Conversely, the ECL6 of proteobacterial BamA presents a ‘hairpin-like’ topology comprising a ‘base’, a ‘stem’, and a ‘terminal loop’, whereas DrBamA’s ECL6 is notably different, lacking the stem and terminal loop (Fig. 3b). In both ‘closed’[8] and ‘open’[11] conformations, the ECL6 of EcBamA consistently adopts a ‘hairpin’ topology that protrudes toward the extracellular side (Fig. 2e and Fig. 3d).
Figure
3.
Analysis of ECL4 and ECL6 in BamA. (a, b) Structural alignments of ECL4s (a) and ECL6s (b) in BamA orthologs from different species. The ECL4 and ECL6 products from the proteobacteria were obtained and colored the same as those in Fig. 1(a). The “hairpin-like” topological organization of EcECL6 is shown in addition to the comparison of the ECL6s. (c) Spot assays in which the chimera EcBamA replaced ECL4 or ECL6 with the corresponding region of DrBamA. (d) Overall structure of the EcBamA barrel domain in the intermediate-open state (PDB: 7TT6) [11]. (e) Polar interactions between residues in the outer helix with other extracellular loops in EcBamA. (f) Spot assays of mutants in the outer helix of EcBamA. (g) Hydrogen bonds of the residues in the stem region of EcECL6. (h) Spot assays of different truncated versions of ECL6 in EcBamA. Each region was deleted by replacement with a GS linker (GSGS for “del 670–706” and GSG for “del 676–699”).
Intrigued by these structural variances, we hypothesized that substituting ECL4 or ECL6 of EcBamA with those from DrBamA might shed light on their roles in OMP folding. Initially, we engineered chimeric versions of EcBamA by replacing its entire ECL4 (Asn538–Asn563) and ECL6 (Leu641–Asn709) with the corresponding sequences from DrBamA (ECL4: Leu614–Thr637; ECL6: Asp727–Thr755), resulting in EcBamA (Dr4) and EcBamA (Dr6), respectively. Furthermore, we combined both modifications to create a double substitution variant, EcBamA (Dr4+Dr6). Western blot analysis revealed that these chimeric EcBamAs were expressed at levels similar to the WT EcBamA in E. coli (Fig. S3b). Spot assays revealed that EcBamA (Dr4+Dr6) significantly impeded cell growth, mirroring the effect observed with the empty vector control (Fig. 3c, line 5). Substitution of either loop individually also led to a remarkable decrease in cell viability by 1–2 orders of magnitude (Fig. 3c, lines 3–4), with ECL4 replacement being more detrimental than ECL6. These results support our hypothesis that extracellular loops are essential for the folding process.
To understand the specific functional impacts of the distinct regions of ECL4 and ECL6, we determined that in EcBamA, the outer helix of ECL4 is characterized by three polar residues (Arg547, Tyr548, and Ser551) forming polar interactions with adjacent loops (ECL7, ECL5, and ECL6) in its open conformation[11] (Fig. 3d, e). In contrast, the outer helix of DrBamA predominantly comprises hydrophobic residues and lacks polar interactions with adjacent loops, which may contribute to its tilted orientation (Fig. 2e) and functional discrepancies. The impaired functionality of EcBamA (Dr4) underscores the potential role of these polar residues in the OMP folding mechanism in E. coli. To further ascertain the significance of these polar residues, we substituted them with hydrophobic alanine (R547A, Y548A, S551A). The mutation significantly compromised the function of EcBamA (Fig. 3f, line 3). Conversely, the function of EcBamA remained relatively stable when adjacent polar residues were mutated to alanine (Q542A, Y550A) (Fig. 3f, line 4). Given that the expression levels of the mutated EcBamA were not significantly affected (Fig. S3c), the results suggest critical roles for specific polar interactions between the outer helix and adjacent ECLs for BamA functionality in E. coli. This conclusion is supported by the lethal effect of mutating Arg547 to alanine observed in a previous study[47]. However, in the distantly related ancient diderm bacterium D. radiodurans, such interactions appear nonessential, suggesting a different, yet to be elucidated, mechanism by which ECL4 contributes to the OMP folding process.
With respect to ECL6, all the BamA orthologs investigated in this study feature a similarly structured ‘base’ (Fig. 3b). Proteobacterial BamAs, however, possess an additional loop structure (Pro670–Val706 in EcBamA), albeit these are less conserved according to multiple sequence alignment (MSA) (Fig. 1b). Structurally, the loop is composed of a ‘stem’ region stabilized with hydrogen bonds and an elongated ‘terminal loop’ connected in between (Fig. 3b and g). To probe the functional importance of different regions of this loop, we engineered various EcBamA truncations, which can be expressed normally in E. coli (Fig. S3d), for spot assays. Notably, removing only the terminal loop residues (Pro676–Leu699) did not significantly affect the functionality of EcBamA (Fig. 3b and h, line 4). However, deleting both the stem and terminal loop (residues 670–706) resulted in severe functional loss, nearly equivalent to the empty vector control (Fig. 3h, line 3). This deficiency was even more pronounced than that of EcBamA, with the entire ECL6 replaced by DrBamA (Fig. 3c, line 4), emphasizing the critical role of the stem region (Pro670–Phe675, Cys700–Val706) in stabilizing ECL6 or assisting in OMP folding in proteobacteria. This finding is consistent with previous findings in which deleting residues 673–702 of BamA was shown to be lethal in E. coli[48]. Intriguingly, DrBamA’s base region contains antiparallel sheets, potentially offering similar ECL6 stabilization functions.
The results of the spot assay combined with the structural comparison demonstrated that ECL4 and ECL6 are critical for the functionality of BamA, particularly the role of the outer helix of ECL4 and the stem region of ECL6 in the folding process of EcBamA and likely other proteobacterial BamAs.
4.
Discussion
In diderm bacteria, the outer membrane is replete with β-barrel proteins (OMPs). Central to assisting their folding is the BAM complex, with BamA serving as the core component. Recent advances, particularly in cryo-EM, have shed light on the mechanism of BamA in facilitating the folding of other OMPs and itself[8–12]. The working model is that POTRA domains (and other BAM components) guide OMP C-terminal β-signals to the β-barrel of BamA, which facilitates folding and insertion into the hydrophobic lipid layer.
D. radiodurans is part of the Deinococcota phylum, which belongs to the larger Terrabacteria clade, alongside other phyla such as Cyanobacteriota, Bacillota, Actinomycetota, Synergistota, Thermotogota, and Chloroflexota. This group includes both diderm and monoderm bacteria, whereas the Gracilicutes clade comprises exclusively diderm bacterial phyla, including Pseudomonadota, the Planctomycetes/Verrucomicrobia/Chlamydiae superphylum, Bacteroidota, and Spirochaetota (Fig. 4)[49]. Recent studies have suggested that these groups diverged following an ancient event characterized by distinct outer membrane tethering systems to the peptidoglycan cell wall[19]. DrBamA and EcBamA exhibit considerable differences: EcBamA features five POTRA domains and four assessor lipoproteins (BamB-E), whereas DrBamA contains six POTRA domains but lacks any identified lipoprotein partners. We propose that an in-depth analysis of BamA structure and function in E. coli and D. radiodurans could offer novel insights into the evolutionary development of bacterial outer membranes. On the basis of the aforementioned differences, it is not surprising that the POTRA domain of EcBamA cannot be replaced with the corresponding region of DrBamA. Some studies have indicated that the β-barrel domain of various proteobacterial BamAs is interchangeable[18], reflecting a possibly conserved folding mechanism. However, our results revealed that the barrel domain of EcBamA cannot be substituted with that of DrBamA, indicating differences between their barrel domains.
To examine the structural differences, we resolved DrBamA’s structure through cryo-EM at a resolution of 3.8 Å, finding that its barrel domain is broadly similar to the 16-strand β-barrel topology of EcBamA but has distinctly arranged extracellular loops, such as ECL4 and ECL6. Substituting these critical loops in EcBamA with those from DrBamA resulted in significant functional impairment, emphasizing their importance in BamA’s function. Specifically, the ECL4 of EcBamA contains a short helix with three polar residues that form key polar interactions with adjacent loops, a feature that is conspicuously absent in DrBamA. Our spot assays confirmed the importance of these interactions in E. coli, alongside the essentiality of the stem region in ECL6, which was also absent in DrBamA.
To further explore the potential connections in outer membrane evolution, we analyzed 24 BamA proteins from eight diderm phyla in Terrabacteria and sixteen from phyla in Gracilicutes, following the categorization by Witwinoski et al.[19] (Fig. 4, Table S3). The structures of these BamAs were either resolved in this study (like DrBamA), obtained from previous studies (such as EcBamA[8], StBamA[46], and NgBamA[48]) or predicted via AlphaFold2[45] (Fig. 3 and Fig. S4). While no clear pattern was evident in the ECL4 loop across the two bacterial clades, our analysis revealed a notable difference in the ECL6 loop. Specifically, 14 of the 16 BamAs examined in Gracilicutes bacteria presented a stem region in their ECL6 loop. In contrast, this feature was found in only two of the eight BamAs from the Terrabacteria clade, as shown in Fig. 4. This distinct structural variation in the ECL6 loop highlights differences between these major bacterial clades in the core OMP folding protein BamA. We speculate that variations in lipids in the outer membrane among species may serve as selection pressures in the evolution of BamA, potentially contributing to the observed structural deviations in ECL6 across different bacterial clades. For example, a notable characteristic of the outer membrane of D. radiodurans is its absence of LPS in the outer leaflet[20], which imparts asymmetry to the outer membrane and influences its fluidity and charge.
Unfortunately, it is unclear whether DrBamA tends to form intrinsic dimers or if the dimeric structure reflects the last stage of BamA folding. More experiments are needed to determine whether DrBamA forms a functional dimer in the native state.
5.
Conclusions
Overall, this study elucidates the crucial roles of BamA’s β-barrel and extracellular loops in OMP folding, revealing significant structural and functional divergences across bacteria. The inability to interchange domains between EcBamA and the evolutionarily distinct DrBamA, alongside the critical importance of specific loop interactions, underscores the evolutionary complexity and specificity of the mechanisms of OMP folding. This research emphasizes the need to broaden our understanding of outer membrane protein folding mechanisms in various microbial organisms, which could unveil novel insights into bacterial physiology and evolution.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (71971201, 72071193, 72188101), the Fundamental Research Funds for the Central Universities (KY2040000049), the Young Taishan Scholars Program of Shandong Province (tsqn202103024), and the Qilu Young Scholars of Shandong University.
Conflict of interest
The authors declare that they have no conflict of interest.
This paper extends the impact of consumer mental accounting on demand to the decisions of a supplier and a retailer, and applies it to wholesale price contracts and revenue sharing contracts.
This paper divides mental accounting by time dimension and portrays the role of mental accounting on consumers’ perception of product valuation and product price by two time coefficients.
This paper reveals the impact of mental accounting on consumers’ perception and purchase decisions, and draws some counterintuitive conclusions.
Kivetz R. Advances in research on mental accounting and reason-based choice. Marketing Letters,1999, 10 (3): 249–266. DOI: 10.1023/A:1008066718905
[3]
Henderson P W, Peterson R A. Mental accounting and categorization. Organizational Behavior and Human Decision Processes,1992, 51 (1): 92–117. DOI: 10.1016/0749-5978(92)90006-S
[4]
Erat S, Bhaskaran S R. Consumer mental accounts and implications to selling base products and add-ons. Marketing Science,2012, 31 (5): 801–818. DOI: 10.1287/mksc.1120.0731
[5]
Prelec D, Loewenstein G. The red and the black: Mental accounting of savings and debt. Marketing Science,1998, 17 (1): 4–28. DOI: 10.1287/mksc.17.1.4
[6]
Gourville J T, Soman D. Payment depreciation: The behavioral effects of temporally separating payments from consumption. Journal of Consumer Research,1998, 25 (2): 160–174. DOI: 10.1086/209533
[7]
Chen L, Kök A G, Tong J D. The effect of payment schemes on inventory decisions: The role of mental accounting. Management Science,2013, 59 (2): 436–451. DOI: 10.1287/mnsc.1120.1638
[8]
Quispe-Torreblanca E G, Stewart N, Gathergood J, et al. The red, the black, and the plastic: Paying down credit card debt for hotels, not sofas. Management Science,2019, 65 (11): 5392–5410. DOI: 10.1287/mnsc.2018.3195
[9]
Dai H, Milkman K L, Riis J. The fresh start effect: Temporal landmarks motivate aspirational behavior. Management Science,2014, 60 (10): 2563–2582. DOI: 10.1287/mnsc.2014.1901
[10]
Ülkü S, Hydock C, Cui S. Making the wait worthwhile: Experiments on the effect of queueing on consumption. Management Science,2020, 66 (3): 1149–1171. DOI: 10.1287/mnsc.2018.3277
[11]
Kaveh A, Nazari M, van der Rest J P, et al. Customer engagement in sales promotion. Marketing Intelligence & Planning,2021, 39 (3): 424–437. DOI: 10.1108/MIP-11-2019-0582
[12]
Ho T H, Png I P L, Reza S. Sunk cost fallacy in driving the world’s costliest cars. Management Science,2018, 64 (4): 1761–1778. DOI: 10.1287/mnsc.2016.2651
[13]
Ho T H, Zhang J. Designing pricing contracts for boundedly rational customers: Does the framing of the fixed fee matter? Management Science,2008, 54 (4): 686–700. DOI: 10.1287/mnsc.1070.0788
[14]
Becker-Peth M, Katok E, Thonemann U W. Designing buyback contracts for irrational but predictable newsvendors. Management Science,2013, 59 (8): 1800–1816. DOI: 10.1287/mnsc.1120.1662
[15]
Davis A M, Katok E, Santamaría N. Push, pull, or both? A behavioral study of how the allocation of inventory risk affects channel efficiency. Management Science,2014, 60 (11): 2666–2683. DOI: 10.1287/mnsc.2014.1940
[16]
Schultz K L, Robinson L W, Thomas L J, et al. The use of framing in inventory decisions. Production and Operations Management,2018, 27 (1): 49–57. DOI: 10.1111/poms.12782
[17]
Gu B, Zhang X. Prospect theory and the newsvendor problem with mental accounting. Journal of Systems Science and Systems Engineering,2020, 29: 525–536. DOI: 10.1007/s11518-019-5445-5
[18]
Shugan S M, Xie J. Advance-selling as a competitive marketing tool. International Journal of Research in Marketing,2005, 22 (3): 351–373. DOI: 10.1016/j.ijresmar.2004.11.004
[19]
Nasiry J, Popescu I. Advance selling when consumers regret. Management Science,2012, 58 (6): 1160–1177. DOI: 10.1287/mnsc.1110.1473
[20]
Yu M, Kapuscinski R, Ahn H S. Advance selling: Effects of interdependent consumer valuations and seller’s capacity. Management Science,2015, 61 (9): 2100–2117. DOI: 10.1287/mnsc.2014.2047
[21]
Ma S, Li G, Sethi S P, et al. Advance selling in the presence of market power and risk-averse consumers. Decision Sciences,2019, 50 (1): 142–169. DOI: 10.1111/deci.12318
[22]
Peng W, Tian Z, Wang Y. Price guarantee for advance selling in the presence of preorder-dependent social learning. International Journal of Production Economics,2020, 219: 115–122. DOI: 10.1016/j.ijpe.2019.05.015
[23]
Zhang Y, Li B, Zhao R. Resale or agency: Pricing strategy for advance selling in a supply chain considering consumers’ loss aversion. IMA Journal of Management Mathematics,2022, 33 (2): 229–254. DOI: 10.1093/imaman/dpab012
[24]
Prasad A, Stecke K E, Zhao X. Advance selling by a newsvendor retailer. Production and Operations Management,2011, 20 (1): 129–142. DOI: 10.1111/j.1937-5956.2010.01133.x
[25]
Wu M, Zhu S X, Teunter R H. Advance selling and advertising: A newsvendor framework. Decision Sciences,2021, 52 (1): 182–215. DOI: 10.1111/deci.12423
[26]
Zhang Z, Lim W, Cui H, et al. Partial refunds as a strategic price commitment device in advance selling in a service industry. European Journal of Operational Research,2021, 291 (3): 1062–1074. DOI: 10.1016/j.ejor.2020.10.001
[27]
Xie J, Liu D, Liang L, et al. Contract choice and advance selling strategy in a supply chain of FAP. PLoS ONE,2022, 17 (3): e0265661. DOI: 10.1371/journal.pone.0265661
[28]
Zhang W, He Y, Gou Q, et al. Optimal advance selling strategy with information provision for omni-channel retailers. Annals of Operations Research,2021: 1–30. DOI: 10.1007/s10479-020-03896-y
He, X., Cheng, W., Fang, Z. et al. Chromium cycle in Red Soil Critical Zone constrained by chromium isotopes. Science of the Total Environment, 2024.
DOI:10.1016/j.scitotenv.2024.176646
2.
Keesari, T.. Tracing the footprints of contaminants in water environment through the application of stable isotopic systematics. Current Opinion in Environmental Science and Health, 2024.
DOI:10.1016/j.coesh.2024.100559
3.
Dietzel, M., Boch, R. Precipitation of CaCO3 in natural and man-made aquatic environments - Mechanisms, analogues, and proxies. Geochemistry, 2024.
DOI:10.1016/j.chemer.2024.126206
Kivetz R. Advances in research on mental accounting and reason-based choice. Marketing Letters,1999, 10 (3): 249–266. DOI: 10.1023/A:1008066718905
[3]
Henderson P W, Peterson R A. Mental accounting and categorization. Organizational Behavior and Human Decision Processes,1992, 51 (1): 92–117. DOI: 10.1016/0749-5978(92)90006-S
[4]
Erat S, Bhaskaran S R. Consumer mental accounts and implications to selling base products and add-ons. Marketing Science,2012, 31 (5): 801–818. DOI: 10.1287/mksc.1120.0731
[5]
Prelec D, Loewenstein G. The red and the black: Mental accounting of savings and debt. Marketing Science,1998, 17 (1): 4–28. DOI: 10.1287/mksc.17.1.4
[6]
Gourville J T, Soman D. Payment depreciation: The behavioral effects of temporally separating payments from consumption. Journal of Consumer Research,1998, 25 (2): 160–174. DOI: 10.1086/209533
[7]
Chen L, Kök A G, Tong J D. The effect of payment schemes on inventory decisions: The role of mental accounting. Management Science,2013, 59 (2): 436–451. DOI: 10.1287/mnsc.1120.1638
[8]
Quispe-Torreblanca E G, Stewart N, Gathergood J, et al. The red, the black, and the plastic: Paying down credit card debt for hotels, not sofas. Management Science,2019, 65 (11): 5392–5410. DOI: 10.1287/mnsc.2018.3195
[9]
Dai H, Milkman K L, Riis J. The fresh start effect: Temporal landmarks motivate aspirational behavior. Management Science,2014, 60 (10): 2563–2582. DOI: 10.1287/mnsc.2014.1901
[10]
Ülkü S, Hydock C, Cui S. Making the wait worthwhile: Experiments on the effect of queueing on consumption. Management Science,2020, 66 (3): 1149–1171. DOI: 10.1287/mnsc.2018.3277
[11]
Kaveh A, Nazari M, van der Rest J P, et al. Customer engagement in sales promotion. Marketing Intelligence & Planning,2021, 39 (3): 424–437. DOI: 10.1108/MIP-11-2019-0582
[12]
Ho T H, Png I P L, Reza S. Sunk cost fallacy in driving the world’s costliest cars. Management Science,2018, 64 (4): 1761–1778. DOI: 10.1287/mnsc.2016.2651
[13]
Ho T H, Zhang J. Designing pricing contracts for boundedly rational customers: Does the framing of the fixed fee matter? Management Science,2008, 54 (4): 686–700. DOI: 10.1287/mnsc.1070.0788
[14]
Becker-Peth M, Katok E, Thonemann U W. Designing buyback contracts for irrational but predictable newsvendors. Management Science,2013, 59 (8): 1800–1816. DOI: 10.1287/mnsc.1120.1662
[15]
Davis A M, Katok E, Santamaría N. Push, pull, or both? A behavioral study of how the allocation of inventory risk affects channel efficiency. Management Science,2014, 60 (11): 2666–2683. DOI: 10.1287/mnsc.2014.1940
[16]
Schultz K L, Robinson L W, Thomas L J, et al. The use of framing in inventory decisions. Production and Operations Management,2018, 27 (1): 49–57. DOI: 10.1111/poms.12782
[17]
Gu B, Zhang X. Prospect theory and the newsvendor problem with mental accounting. Journal of Systems Science and Systems Engineering,2020, 29: 525–536. DOI: 10.1007/s11518-019-5445-5
[18]
Shugan S M, Xie J. Advance-selling as a competitive marketing tool. International Journal of Research in Marketing,2005, 22 (3): 351–373. DOI: 10.1016/j.ijresmar.2004.11.004
[19]
Nasiry J, Popescu I. Advance selling when consumers regret. Management Science,2012, 58 (6): 1160–1177. DOI: 10.1287/mnsc.1110.1473
[20]
Yu M, Kapuscinski R, Ahn H S. Advance selling: Effects of interdependent consumer valuations and seller’s capacity. Management Science,2015, 61 (9): 2100–2117. DOI: 10.1287/mnsc.2014.2047
[21]
Ma S, Li G, Sethi S P, et al. Advance selling in the presence of market power and risk-averse consumers. Decision Sciences,2019, 50 (1): 142–169. DOI: 10.1111/deci.12318
[22]
Peng W, Tian Z, Wang Y. Price guarantee for advance selling in the presence of preorder-dependent social learning. International Journal of Production Economics,2020, 219: 115–122. DOI: 10.1016/j.ijpe.2019.05.015
[23]
Zhang Y, Li B, Zhao R. Resale or agency: Pricing strategy for advance selling in a supply chain considering consumers’ loss aversion. IMA Journal of Management Mathematics,2022, 33 (2): 229–254. DOI: 10.1093/imaman/dpab012
[24]
Prasad A, Stecke K E, Zhao X. Advance selling by a newsvendor retailer. Production and Operations Management,2011, 20 (1): 129–142. DOI: 10.1111/j.1937-5956.2010.01133.x
[25]
Wu M, Zhu S X, Teunter R H. Advance selling and advertising: A newsvendor framework. Decision Sciences,2021, 52 (1): 182–215. DOI: 10.1111/deci.12423
[26]
Zhang Z, Lim W, Cui H, et al. Partial refunds as a strategic price commitment device in advance selling in a service industry. European Journal of Operational Research,2021, 291 (3): 1062–1074. DOI: 10.1016/j.ejor.2020.10.001
[27]
Xie J, Liu D, Liang L, et al. Contract choice and advance selling strategy in a supply chain of FAP. PLoS ONE,2022, 17 (3): e0265661. DOI: 10.1371/journal.pone.0265661
[28]
Zhang W, He Y, Gou Q, et al. Optimal advance selling strategy with information provision for omni-channel retailers. Annals of Operations Research,2021: 1–30. DOI: 10.1007/s10479-020-03896-y
He, X., Cheng, W., Fang, Z. et al. Chromium cycle in Red Soil Critical Zone constrained by chromium isotopes. Science of the Total Environment, 2024.
DOI:10.1016/j.scitotenv.2024.176646
2.
Keesari, T.. Tracing the footprints of contaminants in water environment through the application of stable isotopic systematics. Current Opinion in Environmental Science and Health, 2024.
DOI:10.1016/j.coesh.2024.100559
3.
Dietzel, M., Boch, R. Precipitation of CaCO3 in natural and man-made aquatic environments - Mechanisms, analogues, and proxies. Geochemistry, 2024.
DOI:10.1016/j.chemer.2024.126206



 DownLoad:
DownLoad: