| [1] |
Armitage J P, Schmitt R. Bacterial chemotaxis: Rhodobacter sphaeroides and sinorhizobium meliloti−Variations on a theme? Microbiology, 1997, 12: 3671–3682. doi: 10.1099/00221287-143-12-3671
|
| [2] |
Bren A, Eisenbach M. How signals are heard during bacterial chemotaxis: Protein-protein interactions in sensory signal propagation. Journal of Bacteriology, 2000, 182: 6865–6873. doi: 10.1128/JB.182.24.6865-6873.2000
|
| [3] |
Ud-Din A I M S, Roujeinikova A. Methyl-accepting chemotaxis proteins: A core sensing element in prokaryotes and Archaea. Cellular and Molecular Life Sciences, 2017, 74: 3293–3303. doi: 10.1007/s00018-017-2514-0
|
| [4] |
Wadhams G H, Martin A C, Armitage J P. Identification and localization of a methyl-accepting chemotaxis protein in Rhodobacter sphaeroides. Molecular Microbiology, 2000, 36: 1222–1233. doi: 10.1046/j.1365-2958.2000.01936.x
|
| [5] |
Falke J J, Hazelbauer G L. Transmembrane signaling in bacterial chemoreceptors. Trends in Biochemical Sciences, 2001, 26: 257–265. doi: 10.1016/S0968-0004(00)01770-9
|
| [6] |
Briegel A, Ladinsky M S, Oikonomou C, et al. Structure of bacterial cytoplasmic chemoreceptor arrays and implications for chemotactic signaling. eLife, 2014, 3: 02151. doi: https://doi.org/10.7554/eLife.02151
|
| [7] |
Bi S, Lai L. Bacterial chemoreceptors and chemoeffectors. Cellular and Molecular Life Sciences, 2015, 72: 691–708. doi: 10.1007/s00018-014-1770-5
|
| [8] |
Koler M, Peretz E, Aditya C, et al. Long-term positioning and polar preference of chemoreceptor clusters in E. coli. Nature Communications, 2018, 9: 4444. doi: 10.1038/s41467-018-06835-5
|
| [9] |
Wadhams G H, Armitage J P. Making sense of it all: Bacterial chemotaxis. Nature Reviews Molecular Cell Biology, 2004, 5: 1024–1037. doi: 10.1038/nrm1524
|
| [10] |
Partridge J D, Nhu N T Q, Dufour Y S, et al. Escherichia coli remodels the chemotaxis pathway for swarming. mBio, 2019, 10 (2): e00316-19. doi: https://doi.org/10.1128/mbio.00316-19
|
| [11] |
Welch M, Oosawa K, Aizawa S, et al. Phosphorylation-dependent binding of a signal molecule to the flagellar switch of bacteria. Proceedings of the National Academy of Sciences of the United States of America, 1993, 90: 8787–8791. doi: 10.1073/pnas.90.19.8787
|
| [12] |
Toker A S, MacNab R M. Distinct regions of bacterial flagellar switch protein FliM interact with FliG, FliN and CheY. Journal of Molecular Biology, 1997, 273: 623–634. doi: 10.1006/jmbi.1997.1335
|
| [13] |
Chang Y, Zhang K, Carroll B L, et al. Molecular mechanism for rotational switching of the bacterial flagellar motor. Nature Structural & Molecular Biology, 2020, 27: 1041–1047. doi: https://doi.org/10.1038/s41594-020-0497-2
|
| [14] |
Duke T A J, Bray D. Heightened sensitivity of a lattice of membrane receptors. Proceedings of the National Academy of Sciences of the United States of America, 1999, 96: 10104–10108. doi: 10.1073/pnas.96.18.10104
|
| [15] |
Ames P, Studdert C A, Reiser R H, et al. Collaborative signaling by mixed chemoreceptor teams in Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99: 7060–7065. doi: 10.1073/pnas.092071899
|
| [16] |
Sourjik V, Berg H C. Receptor sensitivity in bacterial chemotaxis. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99: 123–127. doi: 10.1073/pnas.011589998
|
| [17] |
Lamanna A C, Gestwicki J E, Strong L E, et al. Conserved amplification of chemotactic responses through chemoreceptor interactions. Journal of Bacteriology, 2002, 184: 4981–4987. doi: 10.1128/JB.184.18.4981-4987.2002
|
| [18] |
Barkai N, Leibler S. Robustness in simple biochemical networks. Nature, 1997, 387: 913–917. doi: 10.1038/43199
|
| [19] |
Silverman M, Simon M. Chemotaxis in Escherichia coli: Methylation of che gene products. Proceedings of the National Academy of Sciences of the United States of America, 1977, 74: 3317–3321. doi: 10.1073/pnas.74.8.3317
|
| [20] |
Antommattei F M, Munzner J B, Weis R M. Ligand-specific activation of Escherichia coli chemoreceptor transmethylation. Journal of Bacteriology, 2004, 186: 7556–7563. doi: 10.1128/JB.186.22.7556-7563.2004
|
| [21] |
Lan G, Schulmeister S, Sourjik V, et al. Adapt locally and act globally: Strategy to maintain high chemoreceptor sensitivity in complex environments. Molecular Systems Biology, 2011, 7: 475. doi: 10.1038/msb.2011.8
|
| [22] |
Paulick A, Jakovljevic V, Zhang S, et al. Mechanism of bidirectional thermotaxis in Escherichia coli. eLife, 2017, 6: 26607. doi: 10.7554/eLife.26607
|
| [23] |
Meir Y, Jakovljevic V, Oleksiuk O, et al. Precision and kinetics of adaptation in bacterial chemotaxis. Biophysical Journal, 2010, 99: 2766–2774. doi: 10.1016/j.bpj.2010.08.051
|
| [24] |
Micali G, Endres R G. Bacterial chemotaxis: Information processing, thermodynamics, and behavior. Current Opinion in Microbiology, 2016, 30: 8–15. doi: 10.1016/j.mib.2015.12.001
|
| [25] |
Kim C, Jackson M, Lux R, et al. Determinants of chemotactic signal amplification in Escherichia coli. Journal of Molecular Biology, 2001, 307: 119–135. doi: 10.1006/jmbi.2000.4389
|
| [26] |
Hansen C H, Endres R G, Wingreen N S. Chemotaxis in Escherichia coli: A molecular model for robust precise adaptation. PLoS Computational Biology, 2008, 4: e1. doi: 10.1371/journal.pcbi.0040001
|
| [27] |
Li M, Hazelbauer G L. Adaptational assistance in clusters of bacterial chemoreceptors. Molecular Microbiology, 2005, 56: 1617–1626. doi: 10.1111/j.1365-2958.2005.04641.x
|
| [28] |
Endres R G, Wingreen N S. Precise adaptation in bacterial chemotaxis through “assistance neighborhoods”. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103: 13040–13044. doi: 10.1073/pnas.0603101103
|
| [29] |
Mello B A, Tu Y. Perfect and near-perfect adaptation in a model of bacterial chemotaxis. Biophysical Journal, 2003, 84: 2943–2956. doi: 10.1016/S0006-3495(03)70021-6
|
| [30] |
Keymer J E, Endres R G, Skoge M, et al. Chemosensing in Escherichia coli: Two regimes of two-state receptors. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103: 1786–1791. doi: 10.1073/pnas.0507438103
|
| [31] |
Dzinic S H, Shukla M, Mandija I, et al. Variable length tandem repeat polyglutamine sequences in the flexible tether region of the Tsr chemotaxis receptor of Escherichia coli. Microbiology, 2008, 154: 2380–2386. doi: 10.1099/mic.0.2008/016303-0
|
| [32] |
Mello B A, Tu Y. Quantitative modeling of sensitivity in bacterial chemotaxis: The role of coupling among different chemoreceptor species. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100: 8223–8228. doi: 10.1073/pnas.1330839100
|
| [33] |
Lan G, Sartori P, Neumann S, et al. The energy-speed-accuracy trade-off in sensory adaptation. Nature Physics, 2012, 8: 422–428. doi: 10.1038/nphys2276
|
| [34] |
Sourjik V, Vaknin A, Shimizu T S, et al. [17]-in vivo measurement by FRET of pathway activity in bacterial chemotaxis. In: Simon M I, Crane B R, Crane A, editors. Methods in Enzymology: Two-Component Signaling Systems, Part B. Amsterdam: Elsevier, 2007: 365–391.
|
| [35] |
McEvoy M M, Bren A, Eisenbach M, et al. Identification of the binding interfaces on CheY for two of its targets the phosphatase CheZ and the flagellar switch protein FliM. Journal of Molecular Biology, 1999, 289: 1423–1433. doi: 10.1006/jmbi.1999.2830
|
| [36] |
Zhang C, He R, Zhang R, et al. Motor adaptive remodeling speeds up bacterial chemotactic adaptation. Biophysical Journal, 2018, 114: 1225–1231. doi: 10.1016/j.bpj.2018.01.018
|
| [37] |
Liu X, Zhang C, Zhang R, et al. The effect of the second messenger c-di-GMP on bacterial chemotaxis in Escherichia coli. Applied and Environmental Microbiology, 2022, 88: e0037322. doi: 10.1128/aem.00373-22
|
| [38] |
Tu Y, Shimizu T S, Berg H C. Modeling the chemotactic response of Escherichia coli to time-varying stimuli. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105: 14855–14860. doi: 10.1073/pnas.0807569105
|
| [39] |
Jiang L, Ouyang Q, Tu Y. Quantitative modeling of Escherichia coli chemotactic motion in environments varying in space and time. PLoS Computational Biology, 2010, 6: e1000735. doi: 10.1371/journal.pcbi.1000735
|
| [40] |
Monod J, Wyman J, Changeux J P. On the nature of allosteric transitions: A plausible model. Journal of Molecular Biology, 1965, 12: 88–118. doi: 10.1016/S0022-2836(65)80285-6
|
| [41] |
Vladimirov N, Løvdok L, Lebiedz D, et al. Dependence of bacterial chemotaxis on gradient shape and adaptation rate. PLoS Computational Biology, 2008, 4: e1000242. doi: 10.1371/journal.pcbi.1000242
|
| [42] |
Tian M, Zhang C, Zhang R, et al. Collective motion enhances chemotaxis in a two-dimensional bacterial swarm. Biophysical Journal, 2021, 120: 1615–1624. doi: 10.1016/j.bpj.2021.02.021
|
| [43] |
Tu Y, Grinstein G. How white noise generates power-law switching in bacterial flagellar motors. Physical Review Letters, 2005, 94: 208101. doi: 10.1103/PhysRevLett.94.208101
|
| [44] |
Park H, Pontius W, Guet C C, et al. Interdependence of behavioural variability and response to small stimuli in bacteria. Nature, 2010, 468: 819–823. doi: 10.1038/nature09551
|
| [45] |
Zhang C, Zhang R, Yuan J. Growth-dependent behavioral difference in bacterial chemotaxis. Physical Review E, 2017, 95: 062404. doi: 10.1103/PhysRevE.95.062404
|
| [46] |
Cluzel P, Surette M, Leibler S. An ultrasensitive bacterial motor revealed by monitoring signaling proteins in single cells. Science, 2000, 287: 1652–1655. doi: 10.1126/science.287.5458.1652
|
| [47] |
He R, Zhang R, Yuan J. Noise-induced increase of sensitivity in bacterial chemotaxis. Biophysical Journal, 2016, 111: 430–437. doi: 10.1016/j.bpj.2016.06.013
|
| [48] |
Vladimirov N, Lebiedz D, Sourjik V. Predicted auxiliary navigation mechanism of peritrichously flagellated chemotactic bacteria. PLoS Computational Biology, 2010, 6: e1000717. doi: 10.1371/journal.pcbi.1000717
|
| [49] |
Berg H C, Brown D A. Chemotaxis in Escherichia coli analysed by three-dimensional tracking. Nature, 1972, 239: 500–504. doi: 10.1038/239500a0
|
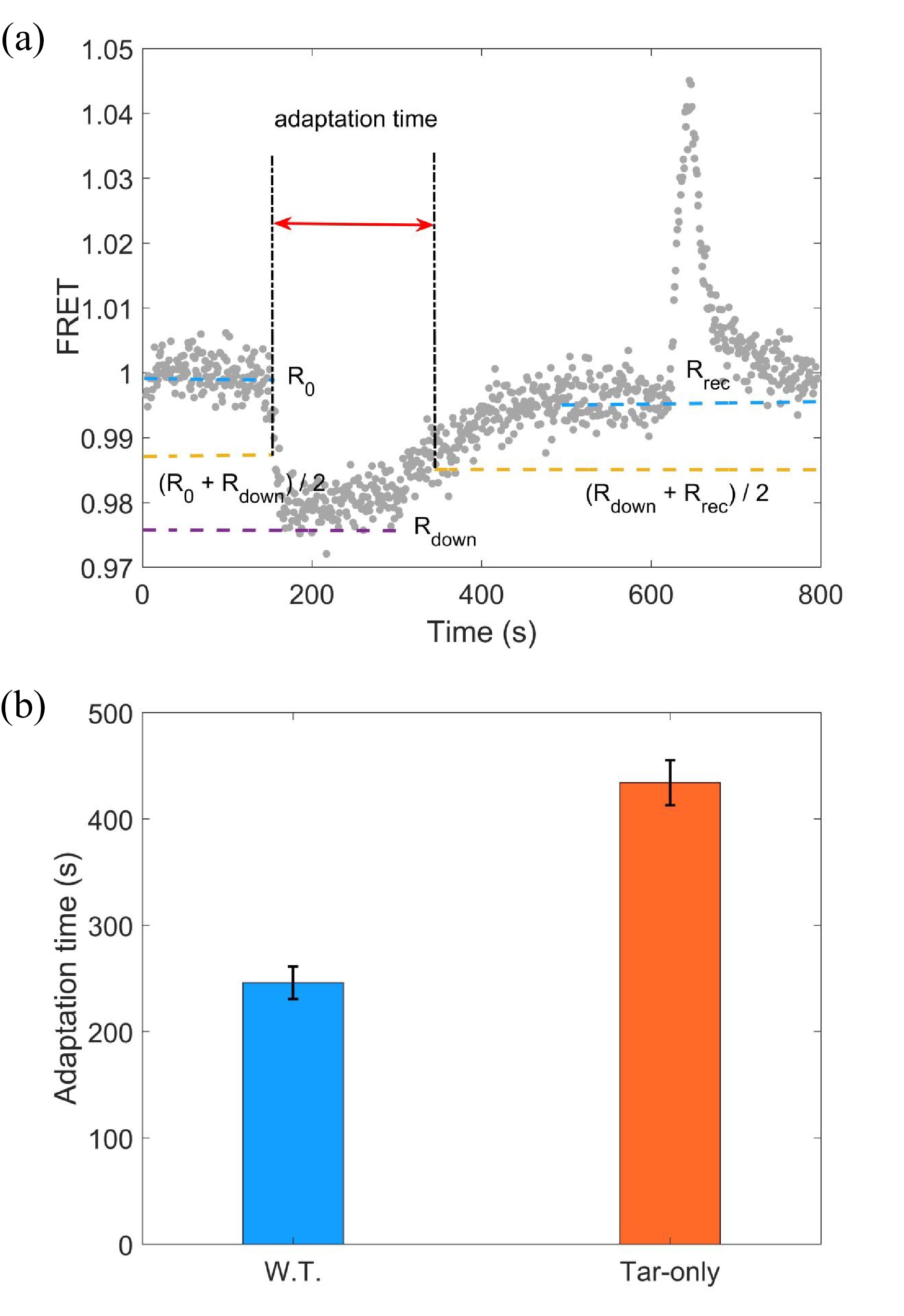
Figure 1. (a) A typical trace of the step response to 10 μmol/L L-aspartate. The stimulus was added at t = 120 s and removed at t = 600 s. The FRET value was defined as the intensity ratio of the YFP to CFP channel. The definition of the adaptation time is indicated. (b) The adaptation time to 10 μmol/L L-aspartate for the wild-type (W.T.) strain (HCB1288-pVS88) and the Tar-only strain (HCB1414-pLC113-pVS88). The errors denote the standard deviations.

Figure 2. (a) The dose‒response measurement to L-aspartate. The blue line is the FRET signal. The red downward arrows denote the time when the stimulus was added, while the black upward arrows indicate the time when the stimulus was removed. The black and red dashed lines represent the pre-stimulus and saturated FRET values, respectively. (b) The dose‒response curve of relative kinase activity (relative to the pre-stimulus value) to the concentration of L-aspartate for both strains. The relative kinase activity was obtained by rescaling the FRET values of 1 (the pre-stimulus value, black dashed line in (a)) to 0.95 (the value after adding a saturated concentration of stimulus, red dashed line in (a)) to the range between 1 and 0. The blue and red lines are the results fitted with the MWC model for the wild-type and mutant strains, respectively. The gray lines indicate the values of relative kinase activity used in the step-response measurements. (c) The relation between adaptation time and relative kinase activity response. The errors denote standard deviations.

Figure 3. (a) The dose‒response curves of the Tar-only strain under 1 μmol/L (red dots, wild-type level) and 0.5 μmol/L (green diamonds) inducer (sodium salicylate). The smooth lines are the results fitted with the MWC model. (b) The relation between adaptation time and relative kinase activity response. The errors denote standard deviations.

Figure 4. (a) Schematic diagram of the bead assay for the flagellar motor. (b) A typical trace of the rotational speed (blue line) of individual motors. The positive and negative values of speed denote CCW and CW rotation, respectively. (c) The CW bias distribution for the wild-type and Tar-only trains. (d) The correlation times for both strains.

Figure 5. (a) Comparison of adaptation times between the wild-type (blue squares) and Tar-only strains (red dots) for all concentrations of L-aspartate. The errors denote standard deviations. (b) The change in mean position over time in a gradient of exponential MeAsp concentration profile L(x) = 91exp(x/5000). The blue dots represent the mean x position. The green squares denote the mean y position. The red line is the linear fit to the blue dots. (c) The effect of adaptation rate on bacterial chemotaxis. Blue and red dots represent the drift velocity in the x direction and the variance of the y positions under different values of kR. The value of kR ranges from 0.005 s−1 to 0.01 s−1 for the wild-type strain, which is marked by the green shaded area.
| [1] |
Armitage J P, Schmitt R. Bacterial chemotaxis: Rhodobacter sphaeroides and sinorhizobium meliloti−Variations on a theme? Microbiology, 1997, 12: 3671–3682. doi: 10.1099/00221287-143-12-3671
|
| [2] |
Bren A, Eisenbach M. How signals are heard during bacterial chemotaxis: Protein-protein interactions in sensory signal propagation. Journal of Bacteriology, 2000, 182: 6865–6873. doi: 10.1128/JB.182.24.6865-6873.2000
|
| [3] |
Ud-Din A I M S, Roujeinikova A. Methyl-accepting chemotaxis proteins: A core sensing element in prokaryotes and Archaea. Cellular and Molecular Life Sciences, 2017, 74: 3293–3303. doi: 10.1007/s00018-017-2514-0
|
| [4] |
Wadhams G H, Martin A C, Armitage J P. Identification and localization of a methyl-accepting chemotaxis protein in Rhodobacter sphaeroides. Molecular Microbiology, 2000, 36: 1222–1233. doi: 10.1046/j.1365-2958.2000.01936.x
|
| [5] |
Falke J J, Hazelbauer G L. Transmembrane signaling in bacterial chemoreceptors. Trends in Biochemical Sciences, 2001, 26: 257–265. doi: 10.1016/S0968-0004(00)01770-9
|
| [6] |
Briegel A, Ladinsky M S, Oikonomou C, et al. Structure of bacterial cytoplasmic chemoreceptor arrays and implications for chemotactic signaling. eLife, 2014, 3: 02151. doi: https://doi.org/10.7554/eLife.02151
|
| [7] |
Bi S, Lai L. Bacterial chemoreceptors and chemoeffectors. Cellular and Molecular Life Sciences, 2015, 72: 691–708. doi: 10.1007/s00018-014-1770-5
|
| [8] |
Koler M, Peretz E, Aditya C, et al. Long-term positioning and polar preference of chemoreceptor clusters in E. coli. Nature Communications, 2018, 9: 4444. doi: 10.1038/s41467-018-06835-5
|
| [9] |
Wadhams G H, Armitage J P. Making sense of it all: Bacterial chemotaxis. Nature Reviews Molecular Cell Biology, 2004, 5: 1024–1037. doi: 10.1038/nrm1524
|
| [10] |
Partridge J D, Nhu N T Q, Dufour Y S, et al. Escherichia coli remodels the chemotaxis pathway for swarming. mBio, 2019, 10 (2): e00316-19. doi: https://doi.org/10.1128/mbio.00316-19
|
| [11] |
Welch M, Oosawa K, Aizawa S, et al. Phosphorylation-dependent binding of a signal molecule to the flagellar switch of bacteria. Proceedings of the National Academy of Sciences of the United States of America, 1993, 90: 8787–8791. doi: 10.1073/pnas.90.19.8787
|
| [12] |
Toker A S, MacNab R M. Distinct regions of bacterial flagellar switch protein FliM interact with FliG, FliN and CheY. Journal of Molecular Biology, 1997, 273: 623–634. doi: 10.1006/jmbi.1997.1335
|
| [13] |
Chang Y, Zhang K, Carroll B L, et al. Molecular mechanism for rotational switching of the bacterial flagellar motor. Nature Structural & Molecular Biology, 2020, 27: 1041–1047. doi: https://doi.org/10.1038/s41594-020-0497-2
|
| [14] |
Duke T A J, Bray D. Heightened sensitivity of a lattice of membrane receptors. Proceedings of the National Academy of Sciences of the United States of America, 1999, 96: 10104–10108. doi: 10.1073/pnas.96.18.10104
|
| [15] |
Ames P, Studdert C A, Reiser R H, et al. Collaborative signaling by mixed chemoreceptor teams in Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99: 7060–7065. doi: 10.1073/pnas.092071899
|
| [16] |
Sourjik V, Berg H C. Receptor sensitivity in bacterial chemotaxis. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99: 123–127. doi: 10.1073/pnas.011589998
|
| [17] |
Lamanna A C, Gestwicki J E, Strong L E, et al. Conserved amplification of chemotactic responses through chemoreceptor interactions. Journal of Bacteriology, 2002, 184: 4981–4987. doi: 10.1128/JB.184.18.4981-4987.2002
|
| [18] |
Barkai N, Leibler S. Robustness in simple biochemical networks. Nature, 1997, 387: 913–917. doi: 10.1038/43199
|
| [19] |
Silverman M, Simon M. Chemotaxis in Escherichia coli: Methylation of che gene products. Proceedings of the National Academy of Sciences of the United States of America, 1977, 74: 3317–3321. doi: 10.1073/pnas.74.8.3317
|
| [20] |
Antommattei F M, Munzner J B, Weis R M. Ligand-specific activation of Escherichia coli chemoreceptor transmethylation. Journal of Bacteriology, 2004, 186: 7556–7563. doi: 10.1128/JB.186.22.7556-7563.2004
|
| [21] |
Lan G, Schulmeister S, Sourjik V, et al. Adapt locally and act globally: Strategy to maintain high chemoreceptor sensitivity in complex environments. Molecular Systems Biology, 2011, 7: 475. doi: 10.1038/msb.2011.8
|
| [22] |
Paulick A, Jakovljevic V, Zhang S, et al. Mechanism of bidirectional thermotaxis in Escherichia coli. eLife, 2017, 6: 26607. doi: 10.7554/eLife.26607
|
| [23] |
Meir Y, Jakovljevic V, Oleksiuk O, et al. Precision and kinetics of adaptation in bacterial chemotaxis. Biophysical Journal, 2010, 99: 2766–2774. doi: 10.1016/j.bpj.2010.08.051
|
| [24] |
Micali G, Endres R G. Bacterial chemotaxis: Information processing, thermodynamics, and behavior. Current Opinion in Microbiology, 2016, 30: 8–15. doi: 10.1016/j.mib.2015.12.001
|
| [25] |
Kim C, Jackson M, Lux R, et al. Determinants of chemotactic signal amplification in Escherichia coli. Journal of Molecular Biology, 2001, 307: 119–135. doi: 10.1006/jmbi.2000.4389
|
| [26] |
Hansen C H, Endres R G, Wingreen N S. Chemotaxis in Escherichia coli: A molecular model for robust precise adaptation. PLoS Computational Biology, 2008, 4: e1. doi: 10.1371/journal.pcbi.0040001
|
| [27] |
Li M, Hazelbauer G L. Adaptational assistance in clusters of bacterial chemoreceptors. Molecular Microbiology, 2005, 56: 1617–1626. doi: 10.1111/j.1365-2958.2005.04641.x
|
| [28] |
Endres R G, Wingreen N S. Precise adaptation in bacterial chemotaxis through “assistance neighborhoods”. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103: 13040–13044. doi: 10.1073/pnas.0603101103
|
| [29] |
Mello B A, Tu Y. Perfect and near-perfect adaptation in a model of bacterial chemotaxis. Biophysical Journal, 2003, 84: 2943–2956. doi: 10.1016/S0006-3495(03)70021-6
|
| [30] |
Keymer J E, Endres R G, Skoge M, et al. Chemosensing in Escherichia coli: Two regimes of two-state receptors. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103: 1786–1791. doi: 10.1073/pnas.0507438103
|
| [31] |
Dzinic S H, Shukla M, Mandija I, et al. Variable length tandem repeat polyglutamine sequences in the flexible tether region of the Tsr chemotaxis receptor of Escherichia coli. Microbiology, 2008, 154: 2380–2386. doi: 10.1099/mic.0.2008/016303-0
|
| [32] |
Mello B A, Tu Y. Quantitative modeling of sensitivity in bacterial chemotaxis: The role of coupling among different chemoreceptor species. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100: 8223–8228. doi: 10.1073/pnas.1330839100
|
| [33] |
Lan G, Sartori P, Neumann S, et al. The energy-speed-accuracy trade-off in sensory adaptation. Nature Physics, 2012, 8: 422–428. doi: 10.1038/nphys2276
|
| [34] |
Sourjik V, Vaknin A, Shimizu T S, et al. [17]-in vivo measurement by FRET of pathway activity in bacterial chemotaxis. In: Simon M I, Crane B R, Crane A, editors. Methods in Enzymology: Two-Component Signaling Systems, Part B. Amsterdam: Elsevier, 2007: 365–391.
|
| [35] |
McEvoy M M, Bren A, Eisenbach M, et al. Identification of the binding interfaces on CheY for two of its targets the phosphatase CheZ and the flagellar switch protein FliM. Journal of Molecular Biology, 1999, 289: 1423–1433. doi: 10.1006/jmbi.1999.2830
|
| [36] |
Zhang C, He R, Zhang R, et al. Motor adaptive remodeling speeds up bacterial chemotactic adaptation. Biophysical Journal, 2018, 114: 1225–1231. doi: 10.1016/j.bpj.2018.01.018
|
| [37] |
Liu X, Zhang C, Zhang R, et al. The effect of the second messenger c-di-GMP on bacterial chemotaxis in Escherichia coli. Applied and Environmental Microbiology, 2022, 88: e0037322. doi: 10.1128/aem.00373-22
|
| [38] |
Tu Y, Shimizu T S, Berg H C. Modeling the chemotactic response of Escherichia coli to time-varying stimuli. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105: 14855–14860. doi: 10.1073/pnas.0807569105
|
| [39] |
Jiang L, Ouyang Q, Tu Y. Quantitative modeling of Escherichia coli chemotactic motion in environments varying in space and time. PLoS Computational Biology, 2010, 6: e1000735. doi: 10.1371/journal.pcbi.1000735
|
| [40] |
Monod J, Wyman J, Changeux J P. On the nature of allosteric transitions: A plausible model. Journal of Molecular Biology, 1965, 12: 88–118. doi: 10.1016/S0022-2836(65)80285-6
|
| [41] |
Vladimirov N, Løvdok L, Lebiedz D, et al. Dependence of bacterial chemotaxis on gradient shape and adaptation rate. PLoS Computational Biology, 2008, 4: e1000242. doi: 10.1371/journal.pcbi.1000242
|
| [42] |
Tian M, Zhang C, Zhang R, et al. Collective motion enhances chemotaxis in a two-dimensional bacterial swarm. Biophysical Journal, 2021, 120: 1615–1624. doi: 10.1016/j.bpj.2021.02.021
|
| [43] |
Tu Y, Grinstein G. How white noise generates power-law switching in bacterial flagellar motors. Physical Review Letters, 2005, 94: 208101. doi: 10.1103/PhysRevLett.94.208101
|
| [44] |
Park H, Pontius W, Guet C C, et al. Interdependence of behavioural variability and response to small stimuli in bacteria. Nature, 2010, 468: 819–823. doi: 10.1038/nature09551
|
| [45] |
Zhang C, Zhang R, Yuan J. Growth-dependent behavioral difference in bacterial chemotaxis. Physical Review E, 2017, 95: 062404. doi: 10.1103/PhysRevE.95.062404
|
| [46] |
Cluzel P, Surette M, Leibler S. An ultrasensitive bacterial motor revealed by monitoring signaling proteins in single cells. Science, 2000, 287: 1652–1655. doi: 10.1126/science.287.5458.1652
|
| [47] |
He R, Zhang R, Yuan J. Noise-induced increase of sensitivity in bacterial chemotaxis. Biophysical Journal, 2016, 111: 430–437. doi: 10.1016/j.bpj.2016.06.013
|
| [48] |
Vladimirov N, Lebiedz D, Sourjik V. Predicted auxiliary navigation mechanism of peritrichously flagellated chemotactic bacteria. PLoS Computational Biology, 2010, 6: e1000717. doi: 10.1371/journal.pcbi.1000717
|
| [49] |
Berg H C, Brown D A. Chemotaxis in Escherichia coli analysed by three-dimensional tracking. Nature, 1972, 239: 500–504. doi: 10.1038/239500a0
|

ISSN 0253-2778
CN 34-1054/N
Copyright © Editorial Office of JUSTC, All Rights Reserved. 皖ICP备05002528号
Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: